Paper Review: Epigenetic Landscape, Cell Differentiation 02
I'll share another review paper about Epigenetic Landscape, it comes from Nature Review, published in 2016.
Transition states and cell fate decisions in epigenetic landscapes [1]
In this paper, they suggest that there might be a fundamental flaw in the reinterpretation of Waddington’s landscape in terms of an assumption of continuous trajectories that branch smoothly at cell fate decisions, and instead propose the notion of a transition state as an alternative that accounts for the gene expression heterogeneity in terms of discontinuities in the mechanisms of fate choice. (Namely, they propose an alternative view: that fate transitions occur in a discontinuous, stochastic manner whereby signals modulate the probability of the transition events.)

Landscapes and dynamical systems
The most famous landscape on the development and evolution of Waddington as follow:
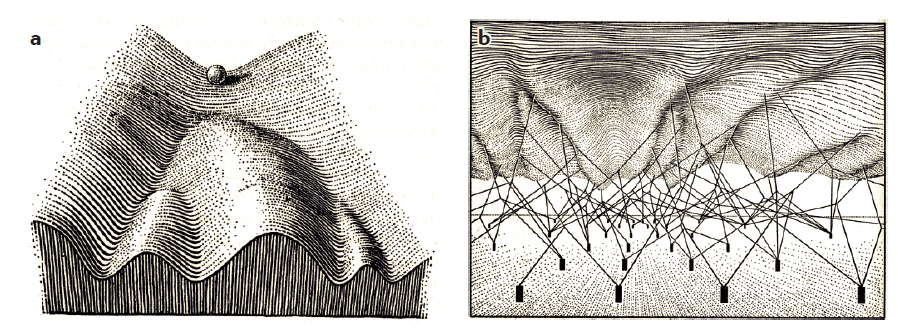
Figure a is famous, and I have explained it in prior articles. Figure b implies that there are genetic regulatory mechanisms responsible for the shape of the landscape.
Any modern interpretation of Waddington’s landscape needs to consider three elements that are built into his vision: the discreteness of the final states or fates, the smooth paths down which cells travel as they move towards these states and the decision events that occur at binary branches in the landscape.
Waddington didn't explicitly point out the dynamical systems theory in the epigenetic landscape, just as a metaphor. Kaufmann uses Boolean networks to formalize the landscape, Huang and colleagues developed this notion further into a multidimensional dynamical systems framework.
However, the true potential landscape is unobtainable and alternatives require some assumptions. The interpretations of the z coordinate in graphical outputs need to be treated with care.
The most critical challenge to Waddington’s vision arises specifically at the branching points in the landscape, where cell fate decisions take place. Different from the smoothness of path in Waddington's theory, alternative theories posit that the initial and final states do not converge and that the decision is discontinuous or that cells must actively ‘jump’ to overcome barriers between discrete fates.
Gene regulatory networks and cell state
The interpretation of sorting genes into "developmental pathways" rests on two important assumptions that permeate gene expression analyses of similar processes: first, given that development proceeds over time, the chains of gene activity must reflect continuous temporal sequences of events; and, second, the process of diversification proceeds through binary choices.
Only high-throughput transcriptional studies are difficult to link to dynamic processes. Alternatively, such data can be used together with perturbation experiments to identify small, regulatory circuits of the system, often arranged in common structural patterns known as network motifs, which are far more amenable to modeling.
The simplest GRN structure that might give rise to a directional state transition event is linear, whereby an input (signaling input) regulates a cascade of downstream genes that results in the equivalent of a ‘gene avalanche’ (some defects). Instead of simplistic linear structures, there is evidence for core feedback loops in regulatory networks that control development, effectively creating circular structures that are adapted to produce stable, discrete cellular states.
Waddington’s potential landscape
It is important to remember that for nonequilibrium, non-gradient systems such as genetic networks, it is not possible to obtain a strict potential function. The alternative is to evaluate the steady-state probabilities of each state or to decompose the vector field and approximate the quasipotential landscape. Then, a value of quasipotential can be calculated for every state that is inversely correlated to the probability that a cell will assume that state, meaning that local ‘low-energy states’ are more densely populated. The changes in the expression profiles correspond to the altering coordinate positions in the landscape.
In the extreme, there are two means by which cells can change their state: by stochastic fluctuations (without requiring any parameter changes) or by extrinsic inputs to the system (alter the parameters of the network). Stochastic fluctuations appear to be an intrinsic feature of some biological systems.
Identifying which model best approximates the biological behavior of cell fate decisions requires experimental evidence of sufficient resolution as to be able to test predictions, ideally using dynamic data from single cells.
Single-cell resolution of fate decisions
The heterogeneity exists in the single-cell level, and the challenge is how to interpret the heterogeneity meaningfully.
Dynamic heterogeneity is functionally relevant to cellular decision making. We need access to the wealth of information harbored within the patterns of these heterogeneities by statistical and computational analyses. (BTW, There are more new methods beyond the following table contains, this is a repo related to single-cell that update in time: https://github.com/seandavi/awesome-single-cell). In these methods, clustering analysis of single-cell data is particularly valuable in identifying very rare or transient populations of cells that might not have been previously observable from low-dimensional or ensemble assays.
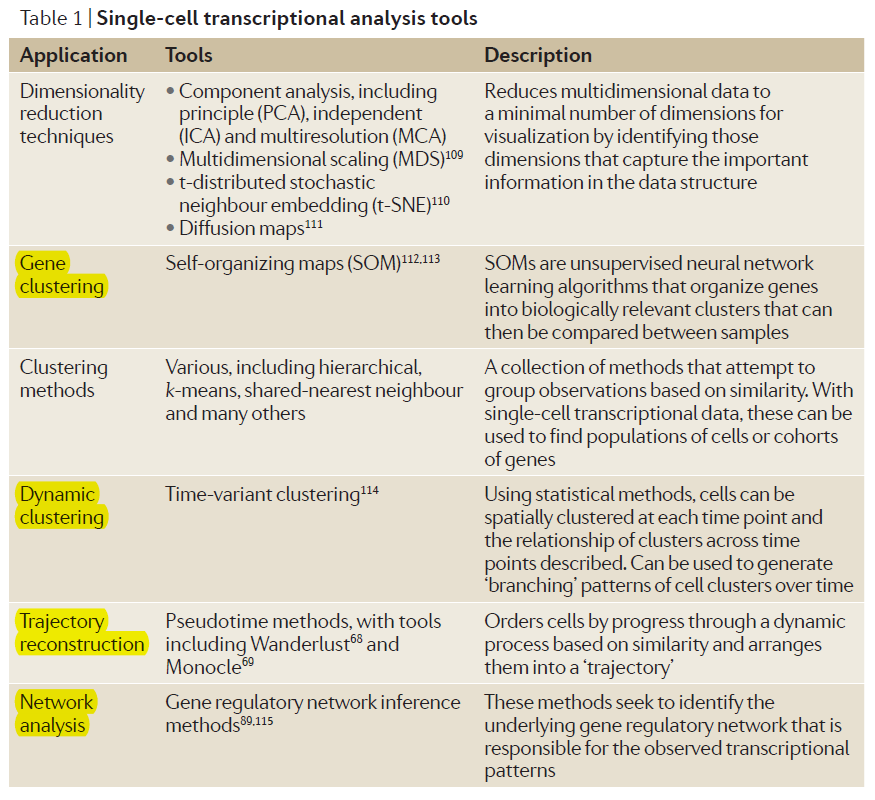
When time-resolved data have been collected (using the snapshots of cells with the same origin to replace a cell at different developmental stages), the major challenge is to extract the salient features in a manner that reveals the underlying dynamic processes. Eg, dimensionality reduction techniques that have been used to create a sequence of ordered events in a dimension referred to as pseudotime.
Under an assumption that influences the pseudotime interpretation — that the transition between states follows a continuous trajectory, which leads to it's unclear that the specific process of cell fate decisions. These mechanisms would be obscured by methods attempting to linearize heterogeneities into a continuous, convergent pathway.
The transition state
- Landscapes and transition states
- A molecular interpretation of the transition state
- The transition state in developmental populations
When cells make a decision, they upregulate the expression levels of the gene cohort of their chosen fate and downregulate those of the alternative one, as has been seen in many systems. An important element in the notion of the transition state is that the passage from one state to another need not be smooth and continuous as portrayed in representations of pseudotime.
The term ‘transition state’ refers to an analogy with the well-known transition state in chemistry, which was introduced to provide a mechanistic underpinning for the progress of a chemical reaction. Similar to the chemical reactions, we surmise that during a fate transition, a GRN can be seen as a reactant that receives a new input, from a signal or from crossing a threshold of its own activity, that leads to either a new network or to a new pattern of connectivity of the existing network. By analogy with a chemical reaction, the cell fate decision process has an intermediate during which the networks explore the state space, and this manifests as heterogeneities in gene expression at the level of single cells. Furthermore, we should notice that the biological transition state is limited by its thermodynamic properties that prevent a formal calculation of potential.
A corollary of the notion of the transition state is that any given phenotypic state might not be associated with just one network state. If GRNs involved in cell fate decisions are hierarchical, it might be that there are key configurations of specific network motifs or small networks that are sufficient to trigger a particular state, independently of additional or downstream gene activity.
< Landscapes and transition states >
Discontinuous cell fate decisions: The actual transitions between states might be discontinuous, and that the observed transcriptional heterogeneity reflects the existence of a dynamic array of transcriptional states, with varied probability of transitions. Then using bifurcation theory to examine the systemic behaviors.
<The transition state in developmental populations>
Although decisions are taken at the level of single cells, at the population level, biases in the bifurcation landscape will result in differences in the number of cells apportioned to each state.
Perspectives
Except for representing data, transition states can frame the role and activity of signals at the decision events, signals represent parameters that can change the structure and the dynamics of the landscape that cells explore. To test these ideas, we shall have to go beyond the analysis of snapshot gene expression data and explore the dynamics of individual genes within individual cells. In the case of a continuous transition, as portrayed by pseudotime, cells at the same point of the fate transition will be transcriptionally very similar to each other, whereas in the case of a discontinuous one, they should be very dissimilar. By observing cells at different stages, according to the continuous pseudotime representation, all sampled intermediates between functional states will follow an identical sequence of cellular states. By contrast, the transition state theory predicts that cells will transition between states through a diverse array of dynamic mechanisms.
Appendix
The bifurcation theories related to cell fate decisions
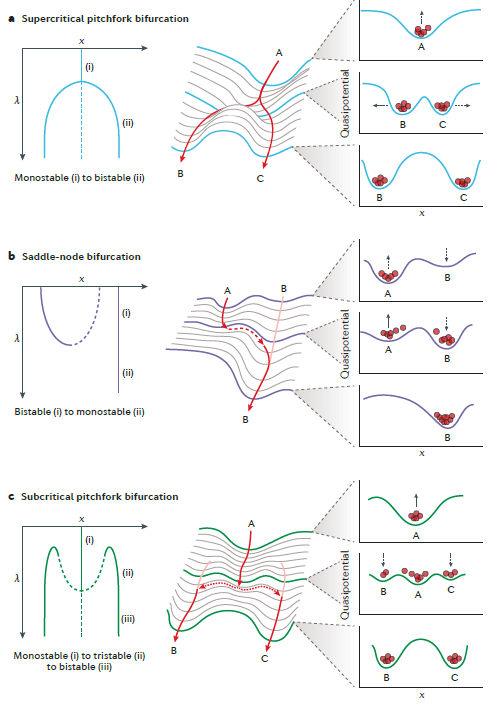
A gene regulatory network (GRN) can be construed as a dynamical system in which the temporal evolution of the system is a function of the parameters of the network. There are several types of bifurcation, each with specific features that produce different behaviors. Here, we focus on three that are pertinent to the study of cell fate decisions. The simplest is the supercritical pitchfork bifurcation, which maps well to the classical Waddington landscape (fate decisions appear smooth and continuous). However, it unable to account for a key feature of cell fate decisions: their irreversibility. In the supercritical pitchfork bifurcation, the temporal dynamics of the landscape depend entirely on the parameter change: if the parameter were reversed, cells from both of the two final states would simply slide back to the initial state. (But the state reversibility is not often observed.)
Alternatively, irreversibility can be achieved through a saddle-node bifurcation, which is proposed in the paper in the last blog I have shared (link to: https://www.cnblogs.com/pear-linzhu/p/12363605.html). The steady states are not created but pre-existed.
The illustration of the transition
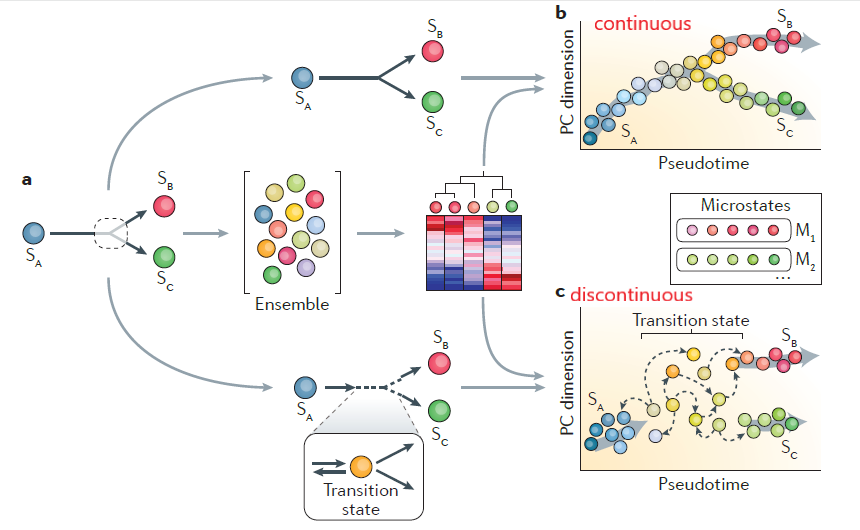
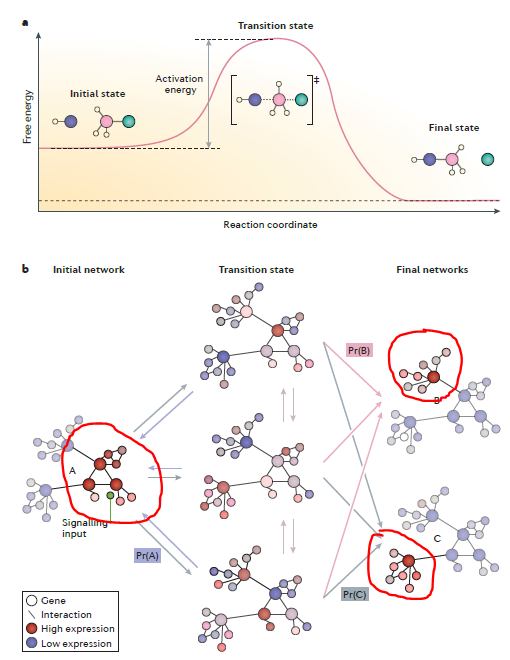
Surmise that the transition state represents a heterogeneous substrate for network selection and fate decisions by factors such as signaling pathways.
Paper Review: Epigenetic Landscape, Cell Differentiation 02的更多相关文章
- Paper Review: Epigenetic Landscape, Cell Differentiation 01
Today, I'll share a review papers about Epigenetic Landscape, the Epigenetic Landscape is related to ...
- (转)CVPR 2016 Visual Tracking Paper Review
CVPR 2016 Visual Tracking Paper Review 本文摘自:http://blog.csdn.net/ben_ben_niao/article/details/52072 ...
- How to transform the day time images to night time ? A series of paper review and some thinkings about this point.
How to transform the day time images to night time ? A series of paper review and some thinkings ab ...
- git review报错一例
在线上修改代码,最后使用git review提交代码审核的时候出现报错如下:[wangshibo@115~]$ vim testfile #修改代码[wangshibo@115~] ...
- lncRNA研究
------------------------------- Long noncoding RNAs are rarely translated in two human cell lines. ( ...
- 基于基因调控网络(Hopfield network)构建沃丁顿表观遗传景观
基因调控网络的概念在之前已经简要介绍过:https://www.cnblogs.com/pear-linzhu/p/12313951.html 沃丁顿表观遗传景观(The Waddington's e ...
- 项目文章|DNA(羟)甲基化研究揭示铁离子依赖表观调控促进狼疮致病性T细胞分化|易基因
易基因(羟)甲基化DNA免疫共沉淀测序(h)MeDIP-seq研究成果见刊<Journal of Clinical Investigation> 2022年5月2日,中南大学湘雅二医院赵明 ...
- single-cell RNA-seq 工具大全
[怪毛匠子-整理] awesome-single-cell List of software packages (and the people developing these methods) fo ...
- Omnibus test
sklearn实战-乳腺癌细胞数据挖掘(博客主亲自录制视频教程) https://study.163.com/course/introduction.htm?courseId=1005269003&a ...
随机推荐
- 51nod:天堂里的游戏
天堂里的游戏 李陶冶 (命题人) System Message (测试) 基准时间限制:1 秒 空间限制:131072 KB 分值: 10 多年后,每当Noder看到吉普赛人,就会想起那个遥远的下午. ...
- TS-Lint 安装方法
1 在 Visual Studio Extensions 里下载 TSLint 2 在项目根目录下放 tslint.json 文件 (里面的规则定死就好 我认为没有特别大的意义)
- JS - 对话框
1,<span onclick="return confirmAct();">执行操作</span> 2, function confirmAct(){ i ...
- 关于linux 交叉编译器的安装
找了几个贴 https://www.cnblogs.com/uestc-mm/p/6656325.html 这个最好
- 指令——mkdir
一个完整的指令的标准格式: Linux通用的格式——#指令主体(空格) [选项](空格) [操作对象] 一个指令可以包含多个选项,操作对象也可以是多个 指令mkdir——(make directory ...
- redis报错MISCONF Redis is configured to save RDB snapshots, but is currently not able to persist
解决方法:通过redis-cli连接到服务器后执行以下命令: config set stop-writes-on-bgsave-error no 注意:这种方法只是忽略了问题,并没有解决问题,具体问题 ...
- 【问题管理】-- Tomcat8部署项目加载静态资源html页面编码错误
1.问题背景及解决方式 最近在回顾Tomcat部署Web项目,自己简单地从Tomcat的下载安装及配置server.xml文件入手,学习Tomcat的项目部署,在自己使用IDEA创建了一个简单地web ...
- enlipse 快捷键
ctrl+shift+o 去掉多余的引用类
- 留学Essay写作中常见的两类要求词盘点
写essay的时候,我们会常常因为各式各样的要求词而头疼:discuss,describing,evaluate,explain,等等,他们之间有何区别?如果你在思考这个问题,那么这篇文章就是为你写的 ...
- 吴裕雄 Bootstrap 前端框架开发——Bootstrap 字体图标(Glyphicons):glyphicon glyphicon-lock
<!DOCTYPE html> <html> <head> <meta charset="utf-8"> <meta name ...